The elucidation of the molecular mechanisms of ion selectivity of hybrid bacterial flagellar motors
The bacterial flagellum acts as the propeller for cell locomotion in a variety of environments. The flagellar motor, consisting of the rotor and the stator, rotates the flagellar filament. All the bacterial flagellar motors characterized so far are energized by either transmembrane electrochemical gradients of protons or sodium ions that are coupled to motility via membrane embedded stator complexes. Some bacteria possess stator complexes of only one ion-coupling type, either protons or sodium, while others have two flagellar systems that have different ion-coupling properties. Use of sodium ions is associated with elevated pH and a high salt environment. Among the reported stators, MotAB-type stators use protons while MotPS- and PomAB-type stators use Na+ as coupling ions. Previously, our group reported that the flagellar motor of Bacillus subtilis is a hybrid motor consisting of both MotAB and MotPS (Ito et al., 2005), and that of Bacillus clausii is a dual-specificity motor consisting of MotAB that has selectivity for both H+ and Na+ (Terahara et al., 2008). We also reported that a novel type of bacterial flagella motor from an alkaliphilic bacterium, Bacillus alcalophilus, can use K+ as a coupling ion (Terahara et al, 2012). It is thought that the flagellar stator of Bacillus species evolved to adjust to the environment. However, the molecular mechanisms of ion selectivity of the flagellar stator have not yet been elucidated. The elucidation of the details will considerably affect the understanding of the operating principles of flagellar motors and the molecular mechanisms of ion selectivity. It will also affect the fields related to the evolution of environmental changes and stresses and nanotechnology. In our project, we conduct researches exploring the differences between a hybrid stator and only one ion-coupling type stator with interdisciplinary collaboration of molecular biology and structure biology, which will enable us to gain a holistic understanding of the mechanisms of ion selectivity of flagellar motors. Furthermore, research will focus on the principal of dynamics of the molecular motor in collaboration with Professor Homma’s group.
Adaptive mechanisms of extreme alkaliphilic microorganisms
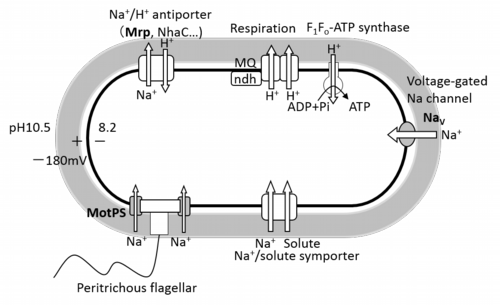
Na+ cycle in alkaliphilic Bacillus
Keywords: Na+/H+ antiporter, Voltage-gated Na+ channel Extreme alkaliphiles, like extremophiles in general, possess numerous structural, metabolic, physiological and bioenergetic adaptations that enable them to function well under their particular “extreme” condition or, in the case of poly-extremophiles, to several extreme conditions at once. If they are facultative extremophiles, many of the adaptations are present even under non-extreme growth conditions. That is, they are “hard-wired” although their expression may increase further under the extreme condition(s). This is presumed to a mechanism that anticipates the need to survive and grow upon a sudden shift to the extreme condition(s). Here, we will summarize a number of different adaptations of alkaliphiles that support their ability to grow optimally at pH values well above 9.0. Some of these species or strains are obligate alkaliphiles that exhibit little or no growth at pH values closer to neutral. Other, facultative alkaliphiles, grow in a range from pH 7.5 to ≥ 11 (Guffanti and Hicks 1991;Yumoto 2007). The facultative alkaliphiles exhibit trade-offs of the type predicted by the “no free lunch” principle defined above. They have a remarkable capacity for growth at pH values much higher than the outer limit of pH 8.5-9.0 for growth of typical neturalophilic bacteria. Facultative alkaliphiles also transition almost seamlessly through a sudden shift from near neutral pH to extremely alkaline pH (Krulwich 1995;Krulwich, et al. 2007;Padan, et al. 2005;Slonczewski, et al. 2009). However, these alkaliphiles exhibit a cost of these remarkable alkaliphilic properties. This deficit is reflected in lower growth rates at near neutral pH than at high pH even though there are greater energy costs for growth at alkaline pH, e.g. pH homeostasis and ATP synthesis (Krulwich, et al. 2007). We suggest that extreme, obligate alkaliphiles represent extreme examples of the “no free lunch” principle in having entirely lost the capacity to grow at neutral pH while excelling at highly alkaline pH values. Each alkaliphile strain that has been examined in some detail displays multiple adaptations that address specific aspects of the challenge of growth at very high pH. For example, they have multiple types of strategies and apparently redundant transporters or enzymes to achieve alkaline pH homeostasis (Padan, et al. 2005;Slonczewski, et al. 2009). This is in accord with a corollary of the “no free lunch” principle which posits that such multiple strategies and their built-in redundancies confer robustness upon the system under the specific condition targeted by the strategies, i.e. survival and growth at alkaline pH in our case. It further predicts that a cost will be exacted in the form of low robustness relative to other organisms under other conditions, i.e., non-alkaline conditions in our case. Thus the “no free lunch” principle is also called the principle of “conservation of fragility” in the engineering literature (Lander, et al. 2009). There follows a set of examples of how this conceptualization of alkaliphily explains data emerging from physiological and bioenergetic studies of several alkaliphiles. We note that a deeper and broader understanding of the design principles underpinning alkaliphily awaits more widespread studies of different extreme alkaliphiles in which mutations can be made in the native setting. Genetically tractable strains are almost entirely unavailable for most extreme alkaliphile types, including extreme Gram-negative alkaliphiles and poly-extremophiles that are robust alkaliphiles that also are thermophilic and/or halophilic (Ma, et al. 2004;Mesbah, et al. 2007;Xue 2006). Without many more genetically tractable and extensively characterized alkaliphile strains, we cannot test the hypotheses about models of robustness and adaptation that are developed from biochemical, genome, transcriptome and proteome data. The “omics” studies can be complemented by studies of mutations of specific molecules in heterologous settings. However, the impact of mutational alterations in specific molecules will often be affected by systems-level of adaptations in the native host in ways that cannot be anticipated by work outside the native setting.
The physiological functions of bacterial voltage-gated Na+ channel, NavBP, in alkaliphilic Bacillus pseudofirmus OF4
Schematic diagram of eukaryotic NaV channel and prokaryotic NaChBac channel.of the page, the caption is also aligned top or bottom, respectively.
■ A voltage-gated sodium channel with homology to the eukaryotic channels that have major roles in physiology and pathophysiology plays a role in alkaliphile pH homeostasis.
■ The NavBP sodium channel of Bacillus pseudofirmus OF4 is required for normal chemotaxis of this alkaliphile and it co-localizes with chemoreceptors at cell poles.